 |
| Iridescent feathers in troodontids - possible? Image by Matt Martyniuk, all rights reserved. (Supreme dino fans may recognize the pose even from this small clip...) |
Sunday, September 8, 2013
Iridescence in Simple Feathers - The Case of the Blue Troodon
Thursday, September 5, 2013
The Tale of the "Sail"
 |
| This version of the painting, "The Fin-Back Lizards" (background: Dimetrodon incisivus, foreground: Naosaurus claviger) by Charles R. Knight, appeared in H.F. Osborn's obituary for E.D. Cope, The Century Magazine (1897). Public domain. |
Sails are often said to be present in other prehistoric animals, like ouranosaurs, spinosaurs, and arizonasaurs, but these are not the quintessential "sails" present in early synapsids like edaphosaurs and dimetrodonts. In the former, the neural spines of the vertebrae are very tall, but also broad and flat, as in normal vertebral columns. These probably anchored muscles, and at the very least supported a ridge of thick soft tissue, not just skin. In dimetrodonts and edaphosaurs*, on the other hand, the neural spines are not just tall, but thin, round, and strut-like. These aren't the kind of vertebrae that would be wrapped in muscle, and may have supported only a thin membrane of skin (I'm not aware of any actual direct evidence for a skin membrane sail, but correct me if I'm wrong).
During the late 1800s, the anatomy and relationships of the sail-backed synapsids was not yet well understood. In a situation weirdly parallel to the famous story about Brontosaurus, the first skeletons of what are now known as Edaphosaurus were found lacking skulls. A small herbivorous skull was actually found first, and given the name Edaphosaurus, but the connection to the sail-backed body was not made until later. The whole saga of Naosaurus, as the headless body was named, was told by Brian Switek at Laelaps. In short, the headless body of Edaphosaurus was seen by E.D. Cope as being very similar to Dimetrodon, and Cope referred a skull to it which is know known to belong to the smooth-spined form rather than the knobby-spined form. When it was discovered that is was actually the small, herbivorous heads already named Edaphosaurus belonged to the headless body, the name Naosaurus was sunk.
 |
| Skull attributed to Naosaurus claviger (but now to Dimetrodon) from Cope 1888, public domain. |
The thing that interested me most about revisiting this story was the whole history of the term "sail" itself. Why were these synapsids referred to as "sail-backs", a term that has since spread to any prehistoric animal with long neural spines?
Many people are aware of the early speculation that sail-backed synapsids used their sails to, well, sail. That is, to literally use their tall dorsal fins to catch the wind and move across water. Most people nowadays also think back on this idea as rather silly. The dorsal fin "sails" were, of course, parallel to the body, like the configuration of a sloop. However, unlike the speculative use of large crests as sails in some pterosaurs, which could at least move the head and neck to change the orientation of the supposed sail, poor dimetrodonts and edaphosaurs would be consigned to getting dragged more or less laterally across the surface of the water. At best, the undulating swimming motion of the torso would cell catch some wind, but the resulting constant change and undulating motion of the sail itself seems like it would make steering very difficult. (Nonetheless, I'm certain I've seen an artistic rendition of this behavior somewhere).
Switek says in his blog post the same thing I and everyone else tend to assume about Cope's sailing hypothesis, which is that Cope suspected "the long spines had a membrane stretched between them and could be used to catch the wind, just like a sail".
But, that's not quite right. It's true that Naosaurus translates as "ship lizard", named for Cope's sail-back hypothesis. But this hypothesis seems to have been specific to Cope's ship lizard, not to the (he thought) closely related Dimetrodon. In fact, if you read some of Cope's original descriptions, you find that the only feature he thought separated Naosaurus from Dimetrodon was the presence of transverse processes on the neural spines. Those are the little thorny side-projections present on the sides of the edaphosaur sail, contrary to the smooth, spike-like bony projections that make up that of dimetrodonts.
Cope thought that it was these side projections, which, while most were broken at the base, he estimated would could be up to half the length of the main neural spine in some specimens, that actually anchored the membranes of the sail! Cope pictured edaphosaurs with a series of membranous sails perpendicular to the torso, not parallel to it, similar to the configuration of the rigging of a large sailing ship rather than a sloop. As Cope said,
"In a full-sized individual, the longest cross-arms, which are the lowest in position, have an expanse of two hundred and sixty millimeters, or ten and a quarter inches, while the spine has about the height of five hundred millimeters (19.75 inches), the body being 60 mm. long. The animal must have presented an extraordinary appearance. Perhaps the yard-arms were connected by membrane with the neural spine or mast, thus serving the animal as a sail, with which he navigated the waters of the Permian lakes." (Cope 1888, p. 294).
While many prehistoric animals are described as having sails, it's interesting to keep in mind that this term seems to first have come about based on a hypothesized structure that was very different from the comparatively "normal" dorsal fins and ridges we're used to seeing today. Cope himself seems to have given up on the idea by the time he was supervising Charles Knight in a restoration of Naosaurus,** which other than the Dimetrodon-like skull, appears relatively normal by modern standards. Still, a proper reconstruction of a truly mast-sailed edaphosaur would be a nice challenge for paleoartists...
*Interestingly, most phylogenetic analyses nowadays suggest that these two types of sail-backed synapsids do not form a natural group with each other. So either the sails evolved convergently, or they are a trait of the common ancestor of the two types of animal. Which would mean our own ancestors were sail-backs!
**Knight later revised his Naosaurus painting, removing the transverse processes altogether and giving it an actual Dimetrodon skull, modifying it into simply a restoration of a Dimetrodon.
References
* Cope, E. D. (1888). Systematic Catalogue of the Species of Vertebrata Found in the Beds of the Permian Epoch in North America with Notes and Descriptions. Transactions of the American Philosophical Society, 16(2), 285-297.
* Osborn, H. F. (1897). A Great Naturalist. Century Magazine, November.
Sunday, September 1, 2013
You're Doing It Wrong : Dino Foot Scales
 |
| Above: Our subject matter. |
These kinds of unjustified assumptions have been rampant in the history of studying stem-birds. Archaeopteryx has traditionally been depicted, incorrectly, with a reversed hallux, and occasionally even with beak-like structures, simply because it's a "bird", and those are features all birds have. Except Archaeopteryx is not a true "bird", it's a stem-bird, more closely related to birds than to any other living animal group, but not a member of the group that includes all modern birds. It's fair to assume that an extinct member of the duck lineage, like Vegavis, had a bill, but that's not necessarily so for, say, Patagopteryx, despite the fact that it is usually referred to as a "bird".
 |
| Modern bird feet, by Philip Henry Gosse, 1849, public domain. Note overlapping scutes on the top surfaces, and pebbly, polygonal reticulae on the bottom surfaces. |
Most paleoartists have absorbed these kinds of warnings, and do a good job of avoiding obvious errors based on typology, the assumption that all species in a certain "type" share "key characteristics." But there are some typological memes in the bird lineage that are more pernicious, possibly because their actual evolution is something most artists don't think about very much.
Take, for example, the bird-like scutes that are almost universally illustrated covering the tarsus (upper foot/lower hind limb) of dinosaurs. Is there any evidence that these were actually present in any given group of non-theropod stem birds? Well... no. Not that I'm aware of (if you know differently, please comment!).
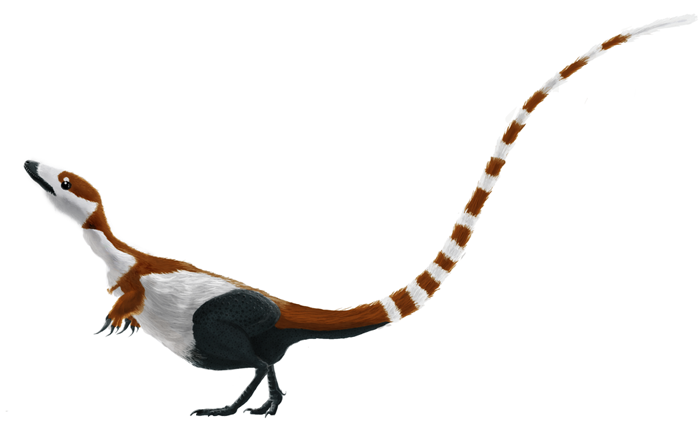 |
| Sinosauropteryx prima with tarsal scutes. Image by Matt Martyniuk, licensed. |
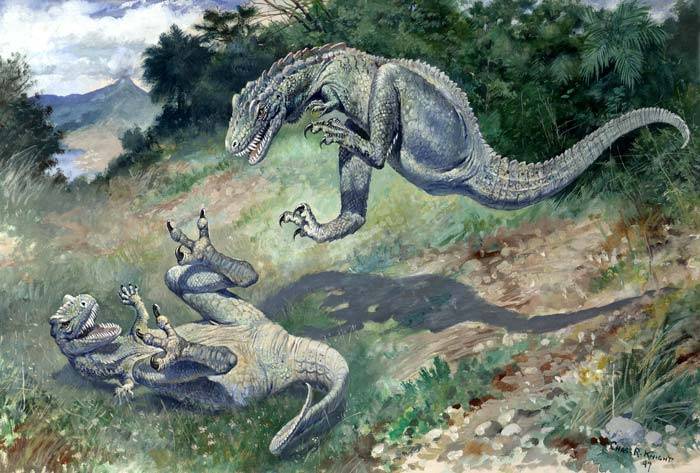{kind=link}
{kind=link}
{kind=link}
Of course, like many paleo-memes that developed during the 1980s, the main idea seems to be using this as a flourish to make otherwise scaly dinosaurs seem more bird-like. And thanks to skin impressions, we know that many dinosaurs had scales, right?
Monday, August 19, 2013
Follow-Up: Judith River Formation = Oldman Formation
In a previous post, I hung my tentative re-identification of the holotype teeth of Deinodon horridus on a rough correlation between the Judith River and Oldman formations, the latter of which is more precisely dated and, more importantly, contains Daspletosaurus torosus, which is a candidate for the owner of Deinodon teeth.
While researching a different topic, I stumbled across a more definitive published correlation of these two formations I wasn't previously aware of. In their 2001 paper on the stratigraphy of the Two Medicine Formation, Horner et al. discuss the correlation of parts of that formation with the Judith River. Horner et al. note that the Judith River can be separated into two basic units divided by a disconformity, corresponding with a marine transgression (when the terrestrial ecosystem was swamped by the rising of the Western Interior Seaway, the sediments deposited by which appear to have been lost in this instance).
Helpfully, Horner et al. note that it is from the lower unit that Hay collected numerous dinosaur teeth which were later described by Leidy as the infamous tooth taxa such as Deinodon, Aublysodon, Trachodon, and Troodon. More helpful still, the paper provides a handy chart showing the arrangement of the strata and including points at which radiometric dates have been taken. The base of the lower Deinodon-bearing unit is dated at about 78 million years old. The next available date is from just above the disconformity (i.e. after the seaway had retreated again) and shows an age of 75.4 million years ago. That's narrowing it down, but there's no date from within the formation from just below the disconformity, which would give us an upper boundary for the Deinodon strata.
But, there's hope. Horner et al. note that Rogers (1998) suggested the disconformity itself probably correlates to around the Willow Creek Anticline in the middle Two Medicine Formation (which contains the famous Egg Mountain Maiasaura nesting site). This segment of the TMF has been dated to 76.7 Ma ago, which may give us a rough upper boundary for the age of Hay's fossil tooth collection.
So, based on this paper at least, it looks like Deinodon and friends were collected from rocks aged somewhere between 78 and 76.7 million years old. Which is about the same age range as the Oldman Formation to the north. So, Deinodon horridus and Daspletosaurus torosus did indeed live at about the same time and in the same region (there were no checkpoints at the US-Canadian border back then!), making it more likely that they represent the same species, and the possibility that Deinodon actually represents Gorgosaurus less likely.
Looks like I'm going to have to create a new tag for Arcane Biostratigraphy and Geology Stuff...
Oh, and somebody in the comments last time asked me to get into Trachodon. This is definitely a subject for a longer blog post, though I'm a bit less excited about it because I'm more pessimistic that it's identity is knowable. But maybe this new info can help us get started. I already mentioned that the Trachodon teeth appear to come from the same strata as Maiasaura (and it's well known that Trachodon's contemporary Troodon formosus is reported from Egg Mountain as well, though T. formosus is also reported from pretty much everywhere and everywhen else...). Could Maiasaura be Trachodon? Perhaps! But it could also be Brachylophosaurus, or maybe even Gryposaurus. And... there's been a rumor going around for a while now that Trachodon teeth are referable to Lambeosaurinae. Which lambeosauines are known from this time and place that could fit the bill? Both Parasaurolophus and Corythosaurus have been reported from the uppermost Oldman, though these may be too young. Hypacrosaurus sp. seems to have been contemporary with Maiasaura, so that could be it...
Yeah, you can see why I'm pessimistic. Tyrannosaurids are rare, and there tend to be only one or two species of tyrannosaurids present in any given ecosystem. Hadrosaurids are... the opposite.
References
Horner, J. R., Schmitt, J. G., Jackson, F., & Hanna, R. (2001). Bones and rocks of the Upper Cretaceous Two Medicine-Judith River clastic wedge complex, Montana. In Field trip guidebook, Society of Vertebrate Paleontology 61st Annual Meeting: Mesozoic and Cenozoic Paleontology in the Western Plains and Rocky Mountains. Museum of the Rockies Occasional Paper (Vol. 3, pp. 3-14).
While researching a different topic, I stumbled across a more definitive published correlation of these two formations I wasn't previously aware of. In their 2001 paper on the stratigraphy of the Two Medicine Formation, Horner et al. discuss the correlation of parts of that formation with the Judith River. Horner et al. note that the Judith River can be separated into two basic units divided by a disconformity, corresponding with a marine transgression (when the terrestrial ecosystem was swamped by the rising of the Western Interior Seaway, the sediments deposited by which appear to have been lost in this instance).
Helpfully, Horner et al. note that it is from the lower unit that Hay collected numerous dinosaur teeth which were later described by Leidy as the infamous tooth taxa such as Deinodon, Aublysodon, Trachodon, and Troodon. More helpful still, the paper provides a handy chart showing the arrangement of the strata and including points at which radiometric dates have been taken. The base of the lower Deinodon-bearing unit is dated at about 78 million years old. The next available date is from just above the disconformity (i.e. after the seaway had retreated again) and shows an age of 75.4 million years ago. That's narrowing it down, but there's no date from within the formation from just below the disconformity, which would give us an upper boundary for the Deinodon strata.
But, there's hope. Horner et al. note that Rogers (1998) suggested the disconformity itself probably correlates to around the Willow Creek Anticline in the middle Two Medicine Formation (which contains the famous Egg Mountain Maiasaura nesting site). This segment of the TMF has been dated to 76.7 Ma ago, which may give us a rough upper boundary for the age of Hay's fossil tooth collection.
So, based on this paper at least, it looks like Deinodon and friends were collected from rocks aged somewhere between 78 and 76.7 million years old. Which is about the same age range as the Oldman Formation to the north. So, Deinodon horridus and Daspletosaurus torosus did indeed live at about the same time and in the same region (there were no checkpoints at the US-Canadian border back then!), making it more likely that they represent the same species, and the possibility that Deinodon actually represents Gorgosaurus less likely.
Looks like I'm going to have to create a new tag for Arcane Biostratigraphy and Geology Stuff...
Oh, and somebody in the comments last time asked me to get into Trachodon. This is definitely a subject for a longer blog post, though I'm a bit less excited about it because I'm more pessimistic that it's identity is knowable. But maybe this new info can help us get started. I already mentioned that the Trachodon teeth appear to come from the same strata as Maiasaura (and it's well known that Trachodon's contemporary Troodon formosus is reported from Egg Mountain as well, though T. formosus is also reported from pretty much everywhere and everywhen else...). Could Maiasaura be Trachodon? Perhaps! But it could also be Brachylophosaurus, or maybe even Gryposaurus. And... there's been a rumor going around for a while now that Trachodon teeth are referable to Lambeosaurinae. Which lambeosauines are known from this time and place that could fit the bill? Both Parasaurolophus and Corythosaurus have been reported from the uppermost Oldman, though these may be too young. Hypacrosaurus sp. seems to have been contemporary with Maiasaura, so that could be it...
Yeah, you can see why I'm pessimistic. Tyrannosaurids are rare, and there tend to be only one or two species of tyrannosaurids present in any given ecosystem. Hadrosaurids are... the opposite.
References
Horner, J. R., Schmitt, J. G., Jackson, F., & Hanna, R. (2001). Bones and rocks of the Upper Cretaceous Two Medicine-Judith River clastic wedge complex, Montana. In Field trip guidebook, Society of Vertebrate Paleontology 61st Annual Meeting: Mesozoic and Cenozoic Paleontology in the Western Plains and Rocky Mountains. Museum of the Rockies Occasional Paper (Vol. 3, pp. 3-14).
Tuesday, August 13, 2013
Did Jurassic Park Name T. rex?
 |
| T. rex illustration by Matt Martyniuk, licensed. |
Sunday, July 14, 2013
Deinodon's Identity Revisited
Tuesday, June 4, 2013
You're Doing It Wrong: CGI Feathered Dinosaurs
 |
| YIKES. Promotional still from When Dinosaur Roamed America, Discovery Channel 2001 |
Saturday, May 11, 2013
The Validity of Lambeosaurus - Anybody Know A Good Lawyer?
 |
| Reconstruction of Didanodon altidens specimen ROM 794 (aka Lambeosaurus lambei, aka Procheneosaurus praeceps) by Matt Martyniuk, all rights reserved. |
The name of one very well-known dinosaur is in such a sorry state that it's like the Manosponylus / Tyrannosaurus debacle squared. The genus Lambeosaurus, a well-known hadrosaurid with a distinctive squared-off crest with a backward-pointed prong, was named twice prior getting its popular moniker, and neither of those names can be considered obsolete, since they were both coined during the 20th century.
The first name given to fossil material (in this case a jaw) now universally attributed to Lambeosaurus was Didanodon. In a 2006 review of hadrosaurs, Lund and Gates stated (without discussion) that the genus and its type species, Didanodon altidens, were nomina nuda, or "naked names" lacking the proper description necessary to establish them. But is this really the case?
Saturday, April 20, 2013
Supporting the Dinosaur/Bird Link in the Era of the MANIAC
I'm posting this more as an open question than a statement of my own opinion, so comments appreciated!
A new paper out in Paleobiology by Verracchio et al. describes the porosity of Troodon formosus eggs and uses the data as evidence to support the hypothesis that troodontids brooded their eggs, like modern birds and other known maniraptorans, rather than burying them, like crocodilians and some other modern birds. In and of itself, this conclusion is interesting in that it pretty much solidifies nest brooding (as opposed to burial) as the ancestral trait for modern birds, and for maniraptorans (or at least chuniaoans*) in general.
I hate to admit it, but my first thought when reading the headline of this news article from PhysOrg was that, yeah, we all assumed that anyway. The unspoken "rule of cool" is that science tends to be more exciting when we find evidence that contradicts previously well-supported hypotheses, rather than confirming hypotheses we all took for granted. Sure, finding the Higgs-Boson was exciting, but not nearly as exciting as not finding it, which could have led to new physics. I assumed, and I'm sure many others did as well, that troodontids brooded their eggs, based on the reasonably secure hypothesis that oviraptorids (which are known to have done so) are more basal. This behavior in troodontids was even depicted nicely in 2011's Dinosaur Revolution. So score another one for phylogenetic bracketing!
(Of course, this is not to imply that all chuniaoans must have brooded their eggs. It's entirely possible that reversals to burial nesting occurred, as with modern megapodes, and this seems especially likely for very large species like some dromaeosaurines. But the odds that any given chuniaoan would not be a brooder are low.)
Aside from all that, the assertion in the PhysOrg headline struck me as particularly meaningless. How could a study of troodontid brooding lend support to the dinosaur/bird hypothesis? This statement would have been accurate a decade ago, but not today. The reason is the moving goalposts of the dinosaur/bird opposition.
A new paper out in Paleobiology by Verracchio et al. describes the porosity of Troodon formosus eggs and uses the data as evidence to support the hypothesis that troodontids brooded their eggs, like modern birds and other known maniraptorans, rather than burying them, like crocodilians and some other modern birds. In and of itself, this conclusion is interesting in that it pretty much solidifies nest brooding (as opposed to burial) as the ancestral trait for modern birds, and for maniraptorans (or at least chuniaoans*) in general.
 |
| Study coauthor Darla Zelenitsky with Troodon formosus nest. Photo by Jay Im, University of Calgary. |
(Of course, this is not to imply that all chuniaoans must have brooded their eggs. It's entirely possible that reversals to burial nesting occurred, as with modern megapodes, and this seems especially likely for very large species like some dromaeosaurines. But the odds that any given chuniaoan would not be a brooder are low.)
 |
| Arctic troodontids, anatomy based on Troodon formosus. Matt Martyniuk, all rights reserved. |
Wednesday, April 10, 2013
What's A Monoclonius?
 |
| Hypothetical restoration of an adult Monoclonius crassus, by Matt Martyniuk. All rights reserved. |
Monoclonius crassus is an iconic ceratopsian, a fixture of many 1970s era dinosaur books, which owes its status almost completely to another species.
I grew up with depictions of Monoclonius in media like Phil Tippet's short film Prehistoric Beast and toys like the DinoRiders figure. Like most representations of Monoclonius in popular culture, these were based on specimens now classified as Centrosaurus apertus (though, actually, that DinoRider looks like it has a genuinely Monoclonius-type frill, long, straight, and unadorned).
{kind=link}
Originally known only from teeth and a fragmentary frill and nasal horn, the Monoclonius was one of the first ceratopsians known to science, found by E.D. Cope in 1876 and named for its configuration of tooth roots ("single sprout" as opposed to the "double sprout" of Diclonius, now known to pertain to a hadrosaur).
Like many of Cope's species, Monoclonius was not recognized for what it really was (a "horned dinosaur") until more complete remains from other ceratopsians likeTriceratops were found by Cope's rival O.C. Marsh. Monoclonius itself remaned enigmatic for many years, though the idea of a Triceratops-like ceratopsian with a single large nasal horn was used by Charles R. Knight in his famous painting of Cope's other dubious ceratopsian, Agathaumas. Knight also incorporated spiny dermal armor associated with some supposed Monoclonius remains, though at least some of this material was later shown to belong to ankylosaurs and pachycephalosaurs.
Monoclonius became iconic when complete skeletons of ceratopsians were found in the Judith River/Dinosaur Park Formation of Montana/Alberta. In the early 1900s, C.H. Sternberg (who had co-discovered the orgiinal Monoclonius fossils with Cope) established that complete specimens classified by Lawrence Lambe as Centrosaurus apertus (and some of which were considered to be Monoclonius and which had already been used to form a picture of that animal) were a distinct species. After this, the genus Monoclonius was dismantled, with former specimens re-assigned to either new or recently established centrosaurine genera.
Matters were complicated by the discovery of the drastic changes centrosaurines went through as they grew, and today the distinctive Monoclonius specimens are generally considered juvenile centrosaurines. Zach Miller has done an awesome rendering of a centrosaurine growth series showing where a traditional "Monoclonius" specimen fits into the sequence.
 |
| Skull of the subadult Monoclonius lowei. Note the three prominent projections at the rear of the parietal frill. Incipient styracosaur-like spikes? |
However, known specimens of Monoclonius aren't a perfect match for juveniles of the contemporary Einiosaurus, as Miller notes. The long, generally flattened frill with larger incipient spikes at the first three positions of the the parietal (Ryan 2006) are reminiscant of Styracosaurus and Einiosaurus, all of about the same geological age. The large size of a specimen sometimes referred to the distinct species Monoclonius lowei compares with some pachyrhinosaurs like the contemporary Achelousaurus, and while these do begin life with a small nasal horn that later develops into a boss, and though it isn't backward-cureved, such drastic changes during ontogony are known in other ceratopsians. It may be that Monoclonius is a juvenile form of (and therefore a senior synonym of) one of these centrosaurines, or it may be a valid species similar to both, possibly a transitional form between centrosaurin-type centrosaurs and pachyrhinosaurs. Ironically, though long mixed up with that genus, Monoclonius doesn't seem to be as great a match for Centrosaurus itself.
In my restoration of a hypothetical, mature Monoclonius (above), I made it generally styracosaur-like, though with more einiosaur-like parietal spikes, and these could alternately be seen as styracosaur parietal spikes which are not yet fully grown. In this way I've tried to hedge my bets: this Monoclonius could either be a mature, intermediate stage between styracosaurs and einiosaurs, or simply an immature but very large styracosaur.
The nasal horn is restored as styracosaur-like as well, a conservative growth trajectory for the shorter, recurved nose horn seen in subadult specimens of M. crassus and M. lowei. However, it's entirely possible that as the nasal horn grew, it swept forward into the hook-like horn of Einiosaurus or even flattened and thickened into the nasal boss seen in Achelousaurus. Both of those pachyrhinosaurs have long parietal spikes like Monoclonius seems to have had, though both only had a single pair jutting from the back of the frill, while Monoclonius seems to have been developing at least three. Though, again, it's possible the transformation was more extreme than I'm assuming for my illustration, and that these incipient parietal horns were resorbed during growth like the epiparietals of chasmosaurines (e.g. Triceratops).
Rather than the centrosaur-like Monoclonius of my childhood, it looks like this fairly plain-looking ceratopsian grew up into something a bit more spectacular. But we'll need further study and, hopefully, more specimens to find out exactly what, and exactly how extreme, that transformation may have been.
* Ryan, M.J. (2006). "The status of the problematic taxon Monoclonius (Ornithischia: Ceratopsidae) and the recognition of adult-sized dinosaur taxa." Geological Society of America Abstracts with Programs, 38(4): 62.
Saturday, April 6, 2013
Friday, March 29, 2013
Who Cares About "Dinosaurs"?
Question: Who cares about dinosaurs?
Short Answer: Marketing departments and monster movie fans.
Long Answer:
This is a philosophical issue that's been on my mind for a while now, inspired by some recent and heated debates over the content of the Dinosaur article at Wikipedia. It also seems to be simmering in the background of a lot of discussions about the recent suggestion that Jurassic Park 4 will not feature modern, scientifically accurate dinosaurians.
Short Answer: Marketing departments and monster movie fans.
 |
| Above: Not what most people think of when you say "dinosaur." (Ashdown Maniraptoran by Matt Martyniuk, all rights reserved). |
This is a philosophical issue that's been on my mind for a while now, inspired by some recent and heated debates over the content of the Dinosaur article at Wikipedia. It also seems to be simmering in the background of a lot of discussions about the recent suggestion that Jurassic Park 4 will not feature modern, scientifically accurate dinosaurians.
Friday, March 22, 2013
No Feathers

As many of you know by now, Jurassic Park 4 director Colin Trevorrow has (basically) announced via tweet that JP4 would not feature feathered dinosaurs but would stick to the 1980s designs of the original film.
Lots of opinions are flying around paleo blogs and they all raise good points. Andrea Cau has been one of the few to defend the decision by noting that changing the dinosaurs would upset the continuity of the films, such as it is (it's not like we're talking about Lord of the Rings style mythology here). Brian Switek has countered that each previous film has completely re-designed the "raptors" anyway with no in-universe explanation. This has even been jarring in the films themselves. At the beginning of the third film, Sam Neil's character Alan Grant has a dream about a raptor, but it's one of the new raptors (with a totally different skull featuring lachrymal horns, new color scheme, and those bizarre psittacosaur-like quills on the neck), not the raptors he or the audience should remember from JP1.
Anyway, it's obvious the producers don't give a hoot about continuity and are trying to appease the JP fans who love the classic dinosaur designs. That's understandable, but it should also be understandable why a significant portion of the fan base is upset. JP is what got many of us into paleontology in the first place, and the first film went out of its way to emphasize the latest science. Steven Spielberg could have gone with dinosaurs the audience expected in 1993: tripodal, tail-dragging, sluggish reptilian beasts. Instead, he made JP the coming out party for the Dinosaur Renaissance, introducing active, bird-like, Bakkerian dinosaurs to a mass audience for the first time in defiance of expectations (dialogue from the film quickly addresses the inaccuracy of the then-classic dinosaurs as well, like noting that the brachiosaur is warm-blooded and doesn't live in swamps).
So it's a bit sad that JP has eaten its own tail and become the self-perpetuating font of inaccurate science the original film was designed to destroy. The franchise is now a slave to its own cannon and the expectations of its audience, rather than having the guts to challenge those expectations. It will inevitably have to rely more and more on cheap built-in loopholes like frog DNA to briefly explain why the dinosaurs are monstrous relics of a bygone era of science less accurate than the CGI characters on a popular children's show.
As Cau noted in his comments, the only thing that could revitalize the series is a reboot. Here's hoping!
Tuesday, January 1, 2013
Double Dewlap! What Does It Mean?
My new book A Field Guide to Mesozoic Birds and Other Winged Dinosaurs is now available! The book can be purchased via Amazon or CreateSpace. A PDF version is available via Lulu (for those of you reading this via RSS, click through to the Web article for handy links on the right side of the post!).
A lot has been made recently about the "new era" dawning in paleoart (or paleontography, or whatever your preferred term is). A move away from the shrink-wrapped and hyper-anatomical artwork that characterized much of the Dinosaur Renaissance. While attention to this new movement has focused mainly on the artwork in the fantastic new book All Yesterdays, Andrea Cau rightfully pointed out on his blog that other artwork has been appearing in this same vein for a while now (though perhaps lacking the publicity it deserves). Cau highlighted the amazing paleo paintings of artist Emiliano Troco, including the rather audacious reconstruction of an Apatosaurus with double dewlap-like structures on the neck.
I guess it's an argument for either the collective consciousness, or that there are in fact a limited number of outlandish yet plausible things you can do when restoring sauropod necks, that I came up with a very similar idea many years ago! My execution of the double dewlap sauropod was somewhat... lacking in technical skill compared to Troco's, but you have to admit it is kind of funny that two people independently came up with such a bizarre idea. After a bit of searching I managed to find a really bad scan of my old sketch, presented below or your amusement!
A lot has been made recently about the "new era" dawning in paleoart (or paleontography, or whatever your preferred term is). A move away from the shrink-wrapped and hyper-anatomical artwork that characterized much of the Dinosaur Renaissance. While attention to this new movement has focused mainly on the artwork in the fantastic new book All Yesterdays, Andrea Cau rightfully pointed out on his blog that other artwork has been appearing in this same vein for a while now (though perhaps lacking the publicity it deserves). Cau highlighted the amazing paleo paintings of artist Emiliano Troco, including the rather audacious reconstruction of an Apatosaurus with double dewlap-like structures on the neck.
 |
| Apatosaurus by Emiliano Troco. |
 |
| My 2002 drawing of an Isisaurus (Titanosaurus colberti at the time) with double dewlaps. Based on a skeletal by Jaime Headden. |
Subscribe to:
Posts (Atom)